Alena J. Markmann1, D. Ryan Bhowmik2,3, Baowei Jiang4, Usaphea P. Vanna5, Michael Van Hoy4,6, Frank Wang4, Yixuan J. Hou7,8, David M. Margolis1,2,9, Ralph S. Baric2,5, Aravinda M. de Silva2, Luther A. Bartelt1,2
1Department of Medicine, Division of Infectious Diseases, University of North Carolina School of Medicine, Chapel Hill, North Carolina
2Department of Microbiology and Immunology, University of North Carolina School of Medicine, Chapel Hill, North Carolina
3Current affiliation: Institute of Bioinformatics, University of Georgia, Athens, Georgia
4BioMedomics Inc., Morrisville, North Carolina
5Department of Pharmacology, University of North Carolina at Chapel Hill, North Carolina
6Current affiliation: Transformative Legal, LLC., Scottsdale, Arizona
7Department of Epidemiology, School of Public Health, University of North Carolina, Chapel Hill, North Carolina
8Current affiliation: Moderna Therapeutics Inc., Cambridge, Massachusetts
9UNC HIV Cure Center, University of North Carolina School of Medicine, Chapel Hill, North Carolina
Alena J. Markmann
Alena.markmann@unchealth.unc.edu
Markmann AJ, Bhowmik DR, Jiang B, Jian B, Vanna UP, Van Hoy M, Wang F, Hou YJ, Margolis DM, Baric RS, de Silva AM, Bartelt LA. A Novel Adaptive Platform for Rapid, Simple Flow-Based Antibody Detection Devices Predicts NAb Levels to SARS-CoV-2. Pathogens and Immunity. 2025;11(1):96-110. doi: 10.20411/pai.v11i1.910
10.20411/pai.v11i1.910
Background: COVID-19 has caused millions of deaths and continues to burden individuals and the healthcare system. Antibodies that neutralize SARS-CoV-2 have proven to be the most reliable markers of immune protection, targets for vaccine development, and approaches for anti-viral antibody-based therapies. Measuring neutralizing antibody (NAb) titers at the bedside could inform individualized shared decision-making with patients regarding the potential benefits of repeating vaccines, use of preventative or therapeutic antibody-based therapies, and, where relevant, collection of COVID-19 convalescent plasma (CCP) with greater efficacy, especially as NAb-escape mutations have guided SARS-CoV-2 variant emergence. However, specific and accessible assays to quantify NAb levels in individuals, including the identification of potential antibody donors at the time of donation, remain unavailable. Therefore, there is a need for platforms that can be rapidly adapted to quantify serum antibody responses with known or expected correlates of protection.
Methods: In this report, we apply a novel semi-quantitative method to an established antibody lateral flow assay (sqLFA) and analyze its ability to detect the presence of functional NAbs in the serum of COVID-19-recovered individuals early in the pandemic.
Results: We found that the sqLFA has a strong positive correlation with the gold-standard microneutralization assay (specificity 80% and sensitivity 90% at a microneutralization cutoff of 1:40).
Conclusions: Taken together, the sqLFA provides a novel point-of-care-based platform for rapid readout of NAb-based immune protection to SARS-CoV-2.
Rapid Diagnostic; COVID-19 Immunity; SARS-CoV-2; Antibody Response; Neutralizing Antibodies; Correlate of Immune Protection
Quantitative antibody detection platforms can assess an individual’s antibody immune response. Rapid and accessible platforms for these assays can be highly valuable at the beginning of a viral epidemic or pandemic for a range of vaccine-preventable infections. Viral infections generate antibody responses, and neutralizing antibody (NAb) responses have been found to be correlates of immune protection (CoP) for many viruses, including SARS-CoV-2 [1]. Despite rapid progress in the SARS-CoV-2 field, the now-normal periodic circulation of novel variants each year and diminishing efficacy of antibody-based therapies have led to the arrested development of rapid point-of-care antibody testing that could help identify those at particular risk of symptomatic infection. These assays can measure various antibody subclasses, including IgG, IgM, and IgA, as well as combinations of these. Clinical lab-based antibody-binding assays, such as enzyme-linked immunosorbent assays (ELISAs) [2], have been developed after being found to provide a satisfactory correlate of protection for some infections such as Hepatitis A and B, Haemophilus influenzae, pertussis, pneumococcus, and tick-borne encephalitis [3]. The most extensively studied antibody immune correlate of protection is for Hepatitis B, where vaccine efficacy studies were performed alongside antibody titer measurements in longitudinal vaccinated cohorts [4]. These studies provide guidance on how antibody CoP could be implemented, but none compare antibody levels to rapid point-of-care testing or provide guidance on the accuracy of rapid antibody testing for CoP.
Lateral flow assays (LFA) are rapid point-of-care antibody-based binding assays that can be performed at home or easily in a clinic, unlike labor-intensive ELISAs, which are usually performed in Clinical Laboratory Improvement Amendent (CLIA)-certified clinical laboratories. Direct neutralization assays are not routinely performed in CLIA-certified clinical laboratories because these labor-intensive assays require BSL3 conditions and are therefore generally restricted to health departments, the Centers for Disease Control and Prevention, or in non-CLIA-certified research laboratories. NAb levels have been identified as a strong correlate of protection for COVID-19, the disease caused by SARS-CoV-2 [2, 5–7]. The immunodominant epitope on the spike envelope protein in SARS-CoV-2 is the receptor-binding domain (RBD), which is the target of 90% of the SARS-CoV-2 NAb response [8]. Antibodies that can bind the RBD can effectively block the virus’s ability to attach to and to enter human epithelial and endothelial cells. In a recent study, researchers found that D614G variant NAbs provided 37% of total protection against delta wave infection at a titer >1:67 [6]. However, NAb responses to natural SARS-CoV-2 infection and vaccination depend on prior variant exposure and are highly variable between individuals in both titer and durability [9–13]. In a recent vaccine efficacy household study with 1461 participants, a NAb titer to Wuhan SARS-CoV-2 strains ≥1:1024 was necessary to confer 92% protection against infection from a delta variant [14]. In a modeling study that used data from 7 vaccine trials and 246 individuals, a NAb titer of 1:54 IU/mL was 50% protective against infection [15]. Clinical trials evaluating antibody-based therapies for COVID-19 have identified a clear benefit to participants who are enrolled prior to antibody seroconversion [16], highlighting the importance that antibody therapies play in early disease.
Additional roles for NAbs as a correlate of clinical outcomes can be drawn from studies of convalescent plasma (CCP). Early in a pandemic, there are often few therapies, and CCP from recovered individuals is often used because it contains antibodies to the novel organism. CCP studies have concluded that it was and continues to be beneficial, especially in immunocompromised populations, if the NAb titer is adequately high (>1:40) and if it is infused early after illness onset [17–21]. However, commercially available rapid antibody detection tests developed during the first year of the pandemic were not rigorously tested against NAb assays to understand whether they correlated with immune protection, and in outcomes-based trials, they were less predictive of CCP efficacy than direct NAb testing [15, 22].
Rapid quantitative antibody assays have dual use: they can be used to rationally design vaccine schedules, particularly for individuals who need repeated vaccination to sustain adequate NAb titers to avoid breakthrough infections, and to identify those who qualify to donate antibody-based therapies in settings where other therapies are unavailable, and in the context of newly emerging variants [13]. For antibody-based assays to inform clinical decisions and public health interventions, quantitative antibody assays need to be developed and validated against more rigorous functional antibody assays, and the technology to detect these antibodies must be widely accessible [23, 24]. Similarly, to progress studies of precise estimates of markers of CoP, the assays or correlations need to be affordable and widely accessible to the community of investigators.
In this study, we use a large cohort of individuals naturally infected with the original SARS-CoV-2 strains and free of interference from prior vaccinations or repeated infections to evaluate a previously clinically validated simple lateral flow-based device [25] in a new prototype reader system that provides a semi-quantitative lateral flow assay (sqLFA), and we correlate the results with the gold-standard NAb titers [26]. We show that our novel platform can be used with a simple lateral flow device to distinguish individuals with high NAb levels from those with lower NAb levels with good sensitivity and specificity. We also find that the sqLFA readout correlates better with NAb levels than other CLIA-certified SARS-CoV-2 antibody assays. This semi-quantitative platform, completed within 20 minutes, can be adapted to various screening antigens rapidly in the setting of a new epidemic or pandemic, as new viral variants emerge, and as CoP data refines rationales for vaccine schedules and antibody-based therapies at the individual patient level.
This study was conducted under Good Clinical Research Practices and was compliant with Institutional Review Board oversight approved by the UNC IRB (20-1141); consent was obtained from all participants. A total of 268 convalescent SARS-CoV-2 plasma samples from patients with natural pre-delta variant infection prior to any vaccination availability were used for this investigation. These samples were collected at a median of 62 days following polymerase chain reaction diagnosis (n = 215) or symptom onset (n = 53), whichever came first, with a range of 12–337 days. Prior characterization of this cohort has been published [11]. Different numbers of the same samples were run in our various antibody assays, and that information is shown in Supplementary Table 1.
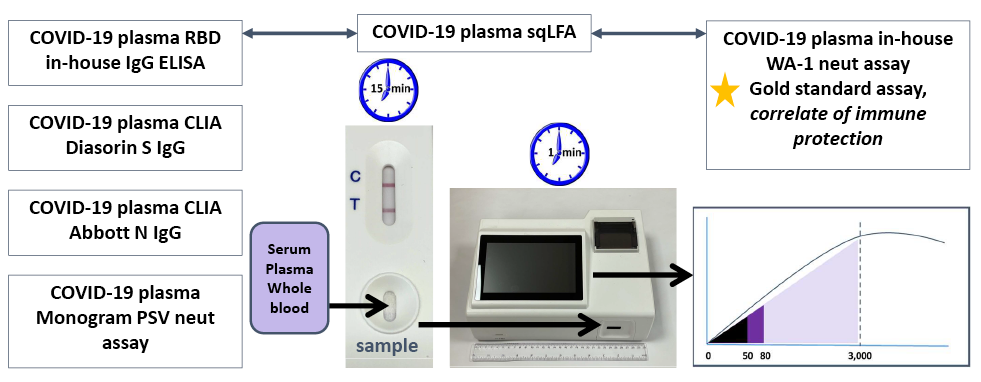
To detect RBD-binding IgG antibodies, we applied a BioMedomics LFA test, which has been separately validated as a rapid diagnostic test to detect SARS-CoV-2 RBD-binding antibodies [25]. The BioMedomics LFA has also been previously validated for whole blood as well as different types of venous samples, including plasma [25]. In the current assay, 10 µL of serum or plasma from each sample was pipetted onto the BioMedomics LFA IgG test strip, followed by 3 drops of buffer solution provided with the kit per manufacturer’s instructions. Each LFA strip was developed for 13 to 15 minutes at room temperature with standard lighting conditions (Figure 1). The LFA strip was then inserted into a prototype reader (Figure 1), which displayed both a qualitative result (Positive/Negative/Intermediate) and a quantitative result in the form of reflective intensity (RI) of gold particles on the LFA strip, which ranged from 0 to 3000. Values were positive according to the manufacturer’s protocol: positive: RI >80, intermediate: RI 50-80, and negative: RI <50. To determine linearity of the RI detector, human IgG antibody was diluted in human serum at different concentrations. Each sample was then tested on the previously validated BioMedomics IgG RBD LFA and read by the prototype reader. The data was then used to create a calibration curve for the detector.
Results from the sqLFA were compared to an in-house SARS-CoV-2 RBD IgG ELISA (end-point-titer), a CLIA-certified lab-run nucleocapsid IgG ELISA (Abbott), a CLIA-certified lab-run full-length Spike IgG ELISA (DiaSorin) [11, 26], and a live virus luciferase reporter-based functional neutralization assay with a readout of 50% neutralization of viral infection (NT50) [26]. The DiaSorin was approved by the Food and Drug Administration (FDA) in 2021 as “Acceptable for Use in the Manufacture of COVID-19 Convalescent Plasma with High Titers of Anti-SARS-CoV-2 Antibodies” at a cutoff of ≥ 87 AU/mL [27]. These assays are listed in Figure 1 and were all performed as previously described [11].
Figure 1. Outline of assays performed. All assays were compared to and correlated with each other as well as with gold-standard assay: WA-1 live virus neutralization assay. Workflow of sqLFA sample application, insertion into prototype reader, and RI readout. 0-50 RI = negative (black), 50-80 = indeterminate (dark purple), >80 = positive (light purple). Abbreviations: RI, reflective intensity; sqLFA, semi-quantitative lateral flow assay.
All statistical analyses and graphs were generated using GraphPad Prism 10.3.1 for Windows [28]. A non-parametric Spearman correlation coefficient was calculated using GraphPad Prism to compare the BioMedomics sqLFA RI to the NT50 titer and other antibody-binding ELISA assays such as our in-house RBD IgG ELISA quantitative end-point titer. All tests were 2-tailed, and a P-value less than 0.05% was considered statistically significant. Receiver operating characteristics (ROC) curves [29] were conducted to determine the sensitivity and specificity of the BioMedomics sqLFA in detection of NT50 at various NAb titer cutoffs. Youden’s J statistic was calculated for each ROC curve and used to maximize both sensitivity and specificity, as presented here. Correlation plots were further analyzed using simple linear regression, with the 95% confidence intervals of the best-fit line shown and shaded in grey.
We found that the sqLFA RI readout for the samples tested ranged from 0 to 2169 and was positively correlated with NT50 titers (Figure 2A), with a Spearman correlation coefficient (rs) of 0.70 (P < 0.0001, n = 268). We also obtained rs for samples that were 14-59 days, ≥ 60 days, or ≥ 90 days post diagnosis or symptom onset; in all cases, rs = 0.70 (P < 0.0001) (data not shown), so within our sample range, the timing post-infection did not change this strong correlation. Among 38 randomly selected samples repeatedly tested on different days (median NT50 titer = 1054, range 0-2076), the intra-assay variability Spearman correlation was rs = 0.94 (P < 0.0001).
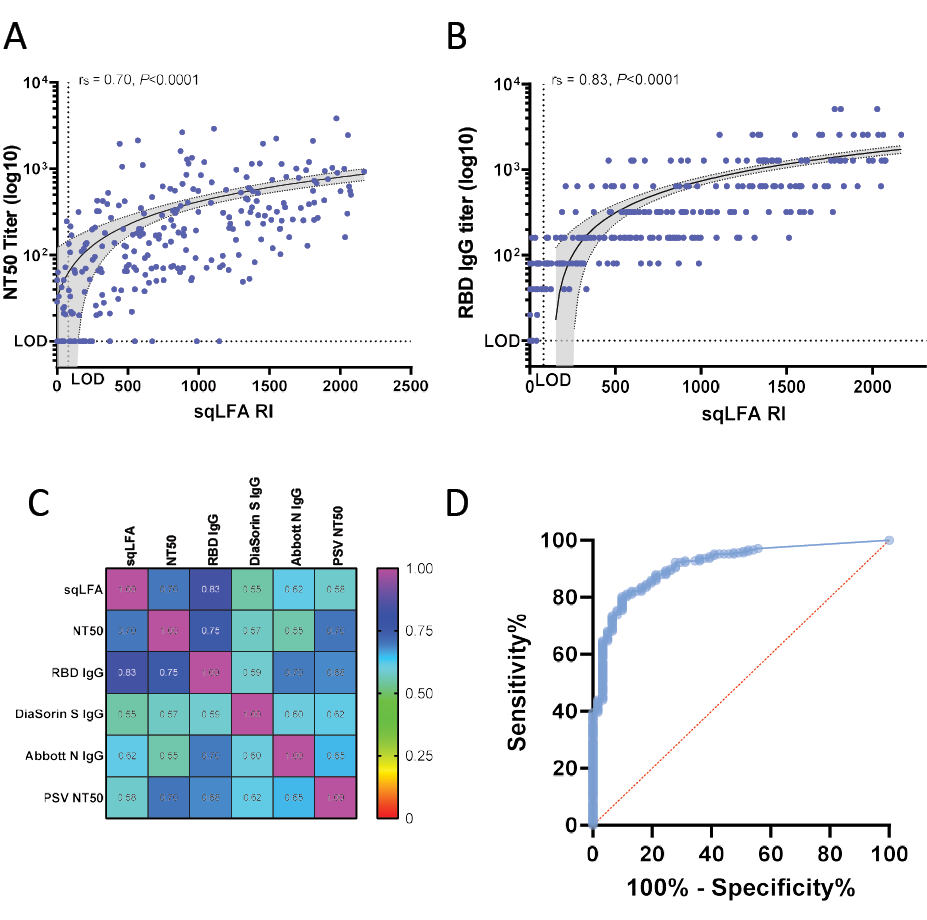
Figure 2. Correlation plots between sqLFA and other serologic assays. A) WA-1 neutralization assays correlation plot with sqLFA. B) RBD IgG in-house ELISA correlation plot with sqLFA, grey shaded area is 95% CI for simple linear regression analysis. C) Heat map of correlation plots for all assays performed with the same samples. D) Example ROC curve of sqLFA performance for all samples with NT50 titer >1:40. Abbreviations: CI, confidence interval; ELISA, enzyme-linked immunosorbent assay; RBD, receptor binding domain; ROC, receiver operating characteristics curve; sqLFA, semi-quantitative lateral flow assay
Our prior work identified that SARS-CoV-2 RBD antibody binding in an in-house assay was a better correlate of NAb NT50 than N-terminal domain, full-length spike protein, and nucleocapsid antigens [11]. Compared to the in-house quantitative RBD IgG ELISA end-point titer data, the sqLFA RI values correlated positively (rs = 0.83; P < 0.0001) (Figure 2B). Among 94 samples tested, the sqLFA correlated modestly with the DiaSorin trimeric spike IgG assay (rs = 0.55; P < 0.0001) (Figure 2C), which is significantly lower than sqLFA vs NT50. Correlation with a commercial SARS-CoV-2 nucleocapsid IgG (Abbott) was even lower at rs = 0.38 (Figure 2C). These findings are consistent with known limitations in both spike and nucleocapsid based IgG binding assays that are not tightly correlated with NT50. Turnaround times for results from these assays were 15 minutes for the sqLFA, 3-6 hours for binding assays (RBD, DiaSorin, Abbott), and a minimum of 72 hours for the live virus NT50.
There is no accepted guideline-directed NAb titer target as a correlate of protection, and consequently, desirable sensitivity and specificity performance beyond diagnosis of exposure is unavailable. We therefore used the literature, including published clinical protocols, to perform analyses using theoretical cutoffs for different NAb titer goals across scenarios. Vaccine breakthrough infection studies, for example, have identified vaccine efficacy and NAb NT50 titer ranges of 1:40, 1:54, and up to 1:1024, depending upon the variant of interest and specific NT50 standard assay used. The original FDA-recommended threshold for therapeutic applications of CCP infusion was NT50 >1:160, and an NT50 >1:640 is an often-cited cutoff for “high titer” serum or plasma samples [30] with predicted greater efficacy.
Multiple ROC analyses were performed to calculate the sensitivity and specificity of the sqLFA RI for detecting samples at different levels of functional Nabs (Table 1). Samples that had detectable NAb titers (NT50 ≥1:40 or other higher cutoffs) were set as positive controls, and those with undetectable NAb titers (or below the cutoff) were set as negative controls using data from our in-house NT50 assay. The area under the curve (AUC) for the sqLFA to detect any NAb NT50 ≥1:40 was 93%, P < 0.0001 (Table 1, Figure 1D). The sensitivity increased to 97% for detecting samples with NT50 ≥1:40 when using the manufacturer’s cutoff of RI > 80. The specificity at this cutoff was low at 46%, which is an expected drop-off at lower titers where functional neutralization is known to be more variable between individuals. The threshold sqLFA cutoff to achieve a specificity >90% for an NT50 ≥1:40 was RI > 457. At an NT50 neutralizing antibody titer of ≥1:54, identified by Khoury et al as a key correlate of 50% protection following vaccination and arguably the most relevant efficacy analysis to date [15], our sqLFA achieved an AUC of 0.85 (P < 0.0001), with both sensitivity and specificity of 82% (Table 1).
We found that at higher NT50 NAb titer cutoffs, the sqLFA had both a high specificity and sensitivity.
Table 1. Sensitivity and Specificity of qLFA for Detecting Neutralizing Antibodies
|
qLFA cutoff |
AUC |
Sensitivity (%) |
95% Cl |
Specificity (%) |
95% Cl |
|
NT50 ≥ 1:40 >457 |
0.93, p<0.0001 |
80 |
74.2–85.0 |
90 |
80.2–95.4 |
|
NT50 ≥ 1:54 >457 |
0.89, p<0.0001 |
82 |
75.5–86.4 |
82 |
72.0–89.3 |
|
NT50 ≥ 1:1024 >872 |
0.71, p=0.0016 |
80 |
58.4–91.9 |
62 |
55.5–67.5 |
ROC analysis results for each NT50 cutoff, showing the sqLFA cutoff with the best Youden’s J statistic to maximize both Sensitivity and Specificity. 95% CI = 95% confidence intervals.
To our knowledge, this is the first study that compares a validated LFA point-of-care test with a semi-quantitative reader platform to a newly published quantitative correlate of protection against SARS-CoV-2 infection, NAb NT50 titers. Here, we demonstrate that the BioMedomics sqLFA rapid platform can serve as a sensitive tool for detecting protective levels of neutralizing antibodies in human samples, providing a proof of concept for future studies and future development of clinically relevant serology-based diagnostics, an area of critical need for respiratory viral infections. From prior work, we know that the immunodominant target on the SARS-CoV-2 spike protein for NAbs is the RBD [2]. The sqLFA platform system with the SARS-CoV-2 RBD antigen has a strong positive correlation with NT50 NAb titers and can be used at various cutoffs to detect different NAb NT50 levels depending on the application. Furthermore, it is easy to use and can be adapted within a few weeks to novel antigens in the event of virus escape mutations or future pandemics, such as the current threat of avian influenza [31]. There is an ongoing need for rapid, deployable tools to assess SARS-CoV-2 immunity post-natural infection or vaccination, especially in immune-susceptible populations. For example, as a future direction, we envision the possibility of generating an LFA with multiple antigens on a single strip to represent current circulating strains as well as vaccine antigens and validating it for use with whole venous blood and potentially fingerstick blood samples.
Our study showed that at an RI > 872, the BioMedomics sqLFA specifically detects NAb levels ≥ 1:1024, which have very recently been shown in 2 separate studies to be a strong CoP against symptomatic alpha or delta strain infection [13, 14, 32]. This correlation far exceeds commercially available spike and nucleocapsid IgG assays available in our clinical laboratory. Using this platform, it is possible to consider a range of patient/individual-directed applications such as timing for repeat COVID-19 vaccine booster (especially in the context of periodic endemic natural infection), need for passive antibody-based therapies in vaccine non-responders, and identification and recruitment of individuals to donate high-titer CCP if such situations arise. The sqLFA point-of-care platform also has a small footprint (Figure 1), low intra-assay variability, and takes less than 20 minutes to set up and obtain a result, making it a great candidate for clinical use and for resource-restricted areas such as much of the rural United States.
Other rapid, semi-quantitative assays have been published and also found to positively correlate with NAb levels, supporting our results here [24, 29, 33, 34]. Two of these assays leverage the interaction between RBD and angiotensin-converting enzyme 2 in their lateral flow development. The COVID-19 Nab-test was studied in a cohort of 79 individuals infected with SARS-CoV-2 and was found to strongly correlate with a microneutralization assay (NT50 ≥1:40), R2 = 0.8 (P < 0.0001), and a reported sensitivity of 79.1% to 99.8% [33]. The quantitative LFA described by Lake et al was compared to neutralization assays from individuals infected and vaccinated and found to have an ROC AUC of 98% at NT50 ≥1:320, with a sensitivity of 90% at an assay cutoff of density units < 263,000 (n = 38 samples) [34]. These assays, like our platform, are based on RBD antigens, show good correlation with NAb titers, and are a promising start in the development of a rapid, quantitative assay surrogate for NAb levels and, therefore, humoral protection against COVID-19 disease. Compared with prior studies, the sqLFA assay reported here used a substantially larger set of samples derived from natural human infection and performed a more granular analysis, reporting sensitivity and specificity across a range of potential NT50 targets.
Since optimal NT50 protective NAb titers as a CoP are still being determined, the present study tested a variety of NT50 cutoffs as a less biased analysis of assay performance.
The main limitation of this type of study is the lack of guidance from the scientific community on what is a good target sensitivity and specificity for a rapid point-of-care test to detect CoP. There has yet to be a longitudinal study evaluating SARS-CoV-2 correlates of protection in a natural infection cohort. In this regard, the lack of widely available functional neutralizing antibody assays may hinder progress in precise immune correlates research and downstream clinical implementation. One could envision a point-of-care assay such as this to enable larger-scale immune protection studies, using a complex assay such as NT50 as a gold standard and establishing targets of protection that can be correlated with the more deployable sqLFA. Although resource limitations prevented inclusion of more recent SARS-CoV-2 variants in this study, recent publications on contemporary strains reveal ongoing strong correlations between RBD-binding antibodies and NAbs [35, 36]. In the event of variant emergence, the sqLFA can be rapidly updated with new variant-specific antigens. Additionally, the sqLFA is expandable to allow inclusion of RBD-targeting IgA and IgM on the LFA strip. Including these antibody isotypes is important as they may contribute to viral neutralization, especially in the first few months post infection, and may further improve sqLFA sensitivity and specificity.
Another limitation is that, to our knowledge, ours is the largest sample size used to study correlations between point-of-care sqLFA results and NT50 titers. However, the cohort may not represent the full range of patient demographics that may benefit from clinical applications of this assay, such as immunocompromised patients who are generally underrepresented (see our prior publication for demographic information on this cohort) [19]. Nonetheless, the relatively low material cost and time investment required to perform the sqLFA point-of-care test make it an attractive adjuvant assay in future larger vaccine efficacy studies that may include NAb measurements to identify a NAb NT50 correlate titer and could simultaneously test the performance of a semi-quantitative point-of-care test such as the BioMedomics sqLFA.
Given ongoing transmission of SARS-CoV-2 variants, as well as known waning antibody levels to both natural infection and vaccination [37], the development of rapid quantitative assays to identify individuals at risk for re-infection is critical. In the SARS-CoV-2 mRNA-vaccinated healthcare worker study by Bergwerk et al, individuals in the cohort with breakthrough infections had a 6- to 7-fold lower mean NAb titer than those who had not experienced a vaccine breakthrough infection [13]. Furthermore, it has been shown that standard 2-dose [38] or even 3-dose [39] mRNA vaccines in solid-organ transplant recipients may not produce an adequate immune response. Current vaccine guidelines recommend annual COVID-19 vaccine boosters for the elderly and those with specific medical conditions or who are immunocompromised [40]. As the landscape of cost-benefit analyses of repeated vaccination for individuals without these major risk factors evolves, having a reliable, easy-to-use, rapid clinical assay that detects protective levels of NAbs and identifies individuals who may need additional SARS-CoV-2 vaccination is a missing yet critical component of current management of SARS-CoV-2 transmission and disease. Readily adaptable antigen-based platforms should be clinically available to be able to evaluate an individual’s degree of vulnerability to disease.
The NIH SeroNet Serocenter of Excellence Award, U54 CA260543, supported generation of laboratory data and the following investigators: AJM, DRB, UPV, YJH, DMM, RSB, AMD, and LAB.
MVH, FW, and BJ are or were employees of BioMedomics Inc. No other conflicts to declare.
Supplementary materials are available at the Pathogens and Immunity website. Supplementary data may be provided by the authors to benefit the reader. Supplementary data are not copyedited and are the sole responsibility of the authors. Questions or comments related to supplementary materials should be addressed to the corresponding author.
Supplementary Table and Figure
1. Escudero-Perez B, Lawrence P, Castillo-Olivares J. Immune Correlates of Protection for Sars-Cov-2, Ebola and Nipah Virus Infection. Front Immunol. 2023;14:1156758. doi: 10.3389/fimmu.2023.1156758. PubMed PMID: 37153606; PMCID: PMC10158532.
2. Piccoli L, Park YJ, Tortorici MA, Czudnochowski N, Walls AC, Beltramello M, Silacci-Fregni C, Pinto D, Rosen LE, Bowen JE, Acton OJ, Jaconi S, Guarino B, Minola A, Zatta F, Sprugasci N, Bassi J, Peter A, De Marco A, Nix JC, Mele F, Jovic S, Rodriguez BF, Gupta SV, Jin F, Piumatti G, Lo Presti G, Pellanda AF, Biggiogero M, Tarkowski M, Pizzuto MS, Cameroni E, Havenar-Daughton C, Smithey M, Hong D, Lepori V, Albanese E, Ceschi A, Bernasconi E, Elzi L, Ferrari P, Garzoni C, Riva A, Snell G, Sallusto F, Fink K, Virgin HW, Lanzavecchia A, Corti D, Veesler D. Mapping Neutralizing and Immunodominant Sites on the Sars-Cov-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell. 2020;183(4):1024-42 e21. doi: 10.1016/j.cell.2020.09.037. PubMed PMID: 32991844; PMCID: PMC7494283.
3. Plotkin SA. Correlates of Protection Induced by Vaccination. Clin Vaccine Immunol. 2010;17(7):1055-65. doi: 10.1128/CVI.00131-10. PubMed PMID: 20463105; PMCID: PMC2897268.
4. Jack AD, Hall AJ, Maine N, Mendy M, Whittle HC. What Level of Hepatitis B Antibody Is Protective? J Infect Dis. 1999;179(2):489-92. doi: 10.1086/314578. PubMed PMID: 9878036.
5. Brady C, Tipton T, Carnell O, Longet S, Gooch K, Hall Y, Salguero J, Tomic A, Carroll M. A Systems Biology Approach to Define Sars-Cov-2 Correlates of Protection. NPJ Vaccines. 2025;10(1):69. doi: 10.1038/s41541-025-01103-2. PubMed PMID: 40229322; PMCID: PMC11997207.
6. Sun K, Bhiman JN, Tempia S, Kleynhans J, Madzorera VS, Mkhize Q, Kaldine H, McMorrow ML, Wolter N, Moyes J, Carrim M, Martinson NA, Kahn K, Lebina L, du Toit JD, Mkhencele T, von Gottberg A, Viboud C, Moore PL, Cohen C, group P-C. Sars-Cov-2 Correlates of Protection from Infection against Variants of Concern. Nat Med. 2024;30(10):2805-12. doi: 10.1038/s41591-024-03131-2. PubMed PMID: 39060660; PMCID: PMC11533127.
7. Goguet E, Olsen CH, Meyer WA, 3rd, Ansari S, Powers JH, 3rd, Conner TL, Coggins SA, Wang W, Wang R, Illinik L, Sanchez Edwards M, Jackson-Thompson BM, Hollis-Perry M, Wang G, Alcorta Y, Wong MA, Saunders D, Mohammed R, Balogun B, Kobi P, Kosh L, Bishop-Lilly K, Cer RZ, Arnold CE, Voegtly LJ, Fitzpatrick M, Luquette AE, Malagon F, Ortega O, Parmelee E, Davies J, Lindrose AR, Haines-Hull H, Moser MS, Samuels EC, Rekedal MS, Graydon EK, Malloy AMW, Tribble DR, Burgess TH, Campbell W, Robinson S, Broder CC, O’Connell RJ, Weiss CD, Pollett S, Laing ED, Mitre E. Immune and Behavioral Correlates of Protection against Symptomatic Post-Vaccination Sars-Cov-2 Infection. Front Immunol. 2024;15:1287504. doi: 10.3389/fimmu.2024.1287504. PubMed PMID: 38566991; PMCID: PMC10985347.
8. Letscher H, Guilligay D, Effantin G, Amen A, Sulbaran G, Burger JA, Bossevot L, Junges L, Leonec M, Morin J, Van Tilbeurgh M, Herate C, Gallouet AS, Relouzat F, van der Werf S, Cavarelli M, Dereuddre-Bosquet N, van Gils MJ, Sanders RW, Poignard P, Le Grand R, Weissenhorn W. Rbd-Depleted Sars-Cov-2 Spike Generates Protective Immunity in Cynomolgus Macaques. NPJ Vaccines. 2025;10(1):63. doi: 10.1038/s41541-025-01113-0. PubMed PMID: 40159504; PMCID: PMC11955555.
9. Crawford KHD, Dingens AS, Eguia R, Wolf CR, Wilcox N, Logue JK, Shuey K, Casto AM, Fiala B, Wrenn S, Pettie D, King NP, Greninger AL, Chu HY, Bloom JD. Dynamics of Neutralizing Antibody Titers in the Months after Severe Acute Respiratory Syndrome Coronavirus 2 Infection. J Infect Dis. 2021;223(2):197-205. doi: 10.1093/infdis/jiaa618. PubMed PMID: 33535236; PMCID: PMC7543487.
10. Long QX, Tang XJ, Shi QL, Li Q, Deng HJ, Yuan J, Hu JL, Xu W, Zhang Y, Lv FJ, Su K, Zhang F, Gong J, Wu B, Liu XM, Li JJ, Qiu JF, Chen J, Huang AL. Clinical and Immunological Assessment of Asymptomatic Sars-Cov-2 Infections. Nat Med. 2020;26(8):1200-4. doi: 10.1038/s41591-020-0965-6. PubMed PMID: 32555424.
11. Markmann AJ, Giallourou N, Bhowmik DR, Hou YJ, Lerner A, Martinez DR, Premkumar L, Root H, van Duin D, Napravnik S, Graham SD, Guerra Q, Raut R, Petropoulos CJ, Wrin T, Cornaby C, Schmitz J, Kuruc J, Weiss S, Park Y, Baric R, de Silva AM, Margolis DM, Bartelt LA. Sex Disparities and Neutralizing-Antibody Durability to Sars-Cov-2 Infection in Convalescent Individuals. mSphere. 2021;6(4):e0027521. doi: 10.1128/mSphere.00275-21. PubMed PMID: 34431693; PMCID: PMC8386415.
12. Roltgen K, Powell AE, Wirz OF, Stevens BA, Hogan CA, Najeeb J, Hunter M, Wang H, Sahoo MK, Huang C, Yamamoto F, Manohar M, Manalac J, Otrelo-Cardoso AR, Pham TD, Rustagi A, Rogers AJ, Shah NH, Blish CA, Cochran JR, Jardetzky TS, Zehnder JL, Wang TT, Narasimhan B, Gombar S, Tibshirani R, Nadeau KC, Kim PS, Pinsky BA, Boyd SD. Defining the Features and Duration of Antibody Responses to Sars-Cov-2 Infection Associated with Disease Severity and Outcome. Sci Immunol. 2020;5(54). doi: 10.1126/sciimmunol.abe0240. PubMed PMID: 33288645; PMCID: PMC7857392.
13. Bergwerk M, Gonen T, Lustig Y, Amit S, Lipsitch M, Cohen C, Mandelboim M, Levin EG, Rubin C, Indenbaum V, Tal I, Zavitan M, Zuckerman N, Bar-Chaim A, Kreiss Y, Regev-Yochay G. Covid-19 Breakthrough Infections in Vaccinated Health Care Workers. N Engl J Med. 2021;385(16):1474-84. doi: 10.1056/NEJMoa2109072. PubMed PMID: 34320281; PMCID: PMC8362591.
14. Regev-Yochay G, Lustig Y, Joseph G, Gilboa M, Barda N, Gens I, Indenbaum V, Halpern O, Katz-Likvornik S, Levin T, Kanaaneh Y, Asraf K, Amit S, Rubin C, Ziv A, Koren R, Mandelboim M, Tokayer NH, Meltzer L, Doolman R, Mendelson E, Alroy-Preis S, Kreiss Y. Correlates of Protection against Covid-19 Infection and Intensity of Symptomatic Disease in Vaccinated Individuals Exposed to Sars-Cov-2 in Households in Israel (Icofs): A Prospective Cohort Study. Lancet Microbe. 2023;4(5):e309-e18. doi: 10.1016/S2666-5247(23)00012-5. PubMed PMID: 36963419; PMCID: PMC10030121.
15. Khoury DS, Cromer D, Reynaldi A, Schlub TE, Wheatley AK, Juno JA, Subbarao K, Kent SJ, Triccas JA, Davenport MP. Neutralizing Antibody Levels Are Highly Predictive of Immune Protection from Symptomatic Sars-Cov-2 Infection. Nat Med. 2021;27(7):1205-11. doi: 10.1038/s41591-021-01377-8. PubMed PMID: 34002089.
16. Group RC. Casirivimab and Imdevimab in Patients Admitted to Hospital with Covid-19 (Recovery): A Randomised, Controlled, Open-Label, Platform Trial. Lancet. 2022;399(10325):665-76. doi: 10.1016/S0140-6736(22)00163-5. PubMed PMID: 35151397; PMCID: PMC8830904.
17. Senefeld JW, Franchini M, Mengoli C, Cruciani M, Zani M, Gorman EK, Focosi D, Casadevall A, Joyner MJ. Covid-19 Convalescent Plasma for the Treatment of Immunocompromised Patients: A Systematic Review and Meta-Analysis. JAMA Netw Open. 2023;6(1):e2250647. doi: 10.1001/jamanetworkopen.2022.50647. PubMed PMID: 36633846; PMCID: PMC9857047.
18. Hoffmann S, Schrezenmeier E, Desmarets M, Halleck F, Durrbach A, Peters L, Tremmel AT, Seidel A, Fuhrer M, Bachmann F, Schrezenmeier J, Greiner J, Korper S, Hofmann H, Ludwig C, Vieweg C, Jahrsdorfer B, Budde K, Schmidt M, Munch J, Joher N, Daguindau E, Gruner B, Brunotte G, Vauchy C, Seifried E, Bradshaw D, Estcourt LJ, Roberts DJ, Toussirot E, Rijnders B, Tiberghien P, Schrezenmeier H. Early, Very High-Titre Convalescent Plasma Therapy in Clinically Vulnerable Individuals with Mild Covid-19: An International, Randomised, Open-Label Trial. EBioMedicine. 2025;113:105613. doi: 10.1016/j.ebiom.2025.105613. PubMed PMID: 40020259; PMCID: PMC11919330.
19. Bartelt LA, Markmann AJ, Nelson B, Keys J, Root H, Henderson HI, Kuruc J, Baker C, Bhowmik DR, Hou YJ, Premkumar L, Cornaby C, Schmitz JL, Weiss S, Park Y, Baric R, de Silva AM, Lachiewicz A, Napravnik S, van Duin D, Margolis DM. Outcomes of Convalescent Plasma with Defined High Versus Lower Neutralizing Antibody Titers against Sars-Cov-2 among Hospitalized Patients: Coronavirus Inactivating Plasma (Covip) Study. mBio. 2022;13(5):e0175122. doi: 10.1128/mbio.01751-22. PubMed PMID: 36135380; PMCID: PMC9601237.
20. Denkinger CM, Janssen M, Schakel U, Gall J, Leo A, Stelmach P, Weber SF, Krisam J, Baumann L, Stermann J, Merle U, Weigand MA, Nusshag C, Bullinger L, Schrezenmeier JF, Bornhauser M, Alakel N, Witzke O, Wolf T, Vehreschild M, Schmiedel S, Addo MM, Herth F, Kreuter M, Tepasse PR, Hertenstein B, Hanel M, Morgner A, Kiehl M, Hopfer O, Wattad MA, Schimanski CC, Celik C, Pohle T, Ruhe M, Kern WV, Schmitt A, Lorenz HM, Souto-Carneiro M, Gaeddert M, Halama N, Meuer S, Krausslich HG, Muller B, Schnitzler P, Parthe S, Bartenschlager R, Gronkowski M, Klemmer J, Schmitt M, Dreger P, Kriegsmann K, Schlenk RF, Muller-Tidow C. Anti-Sars-Cov-2 Antibody-Containing Plasma Improves Outcome in Patients with Hematologic or Solid Cancer and Severe Covid-19: A Randomized Clinical Trial. Nat Cancer. 2023;4(1):96-107. doi: 10.1038/s43018-022-00503-w. PubMed PMID: 36581734; PMCID: PMC9886549.
21. Lacombe K, Hueso T, Porcher R, Mekinian A, Chiarabini T, Georgin-Lavialle S, Ader F, Saison J, Martin-Blondel G, De Castro N, Bonnet F, Cazanave C, Francois A, Morel P, Hermine O, Pourcher V, Michel M, Lescure X, Soussi N, Brun P, Pommeret F, Sellier P, Rousset S, Piroth L, Michot JM, Baron G, de Lamballerie X, Mariette X, Tharaux PL, Resche-Rigon M, Ravaud P, Simon T, Tiberghien P. Use of Covid-19 Convalescent Plasma to Treat Patients Admitted to Hospital for Covid-19 with or without Underlying Immunodeficiency: Open Label, Randomised Clinical Trial. BMJ Med. 2023;2(1):e000427. doi: 10.1136/bmjmed-2022-000427. PubMed PMID: 37920150; PMCID: PMC10619082.
22. Abbasi J. The Flawed Science of Antibody Testing for Sars-Cov-2 Immunity. JAMA. 2021;326(18):1781-2. doi: 10.1001/jama.2021.18919. PubMed PMID: 34673883.
23. Gundlapalli AV, Salerno RM, Brooks JT, Averhoff F, Petersen LR, McDonald LC, Iademarco MF, Response CC-. Sars-Cov-2 Serologic Assay Needs for the Next Phase of the Us Covid-19 Pandemic Response. Open Forum Infect Dis. 2021;8(1):ofaa555. doi: 10.1093/ofid/ofaa555. PubMed PMID: 33442555; PMCID: PMC7717402.
24. Mallory M, Munt JE, Narowski TM, Castillo I, Cuadra E, Pisanic N, Fields P, Powers JM, Dickson A, Harris R, Wargowsky R, Moran S, Allabban A, Raphel K, McCaffrey TA, Brien JD, Heaney CD, Lafleur JE, Baric RS, Premkumar L. Covid-19 Point-of-Care Tests Can Identify Low-Antibody Individuals: In-Depth Immunoanalysis of Boosting Benefits in a Healthy Cohort. Sci Adv. 2024;10(24):eadi1379. doi: 10.1126/sciadv.adi1379. PubMed PMID: 38865463; PMCID: PMC11168476.
25. Li Z, Yi Y, Luo X, Xiong N, Liu Y, Li S, Sun R, Wang Y, Hu B, Chen W, Zhang Y, Wang J, Huang B, Lin Y, Yang J, Cai W, Wang X, Cheng J, Chen Z, Sun K, Pan W, Zhan Z, Chen L, Ye F. Development and Clinical Application of a Rapid Igm-Igg Combined Antibody Test for Sars-Cov-2 Infection Diagnosis. J Med Virol. 2020;92(9):1518-24. doi: 10.1002/jmv.25727. PubMed PMID: 32104917; PMCID: PMC7228300.
26. Premkumar L, Segovia-Chumbez B, Jadi R, Martinez DR, Raut R, Markmann A, Cornaby C, Bartelt L, Weiss S, Park Y, Edwards CE, Weimer E, Scherer EM, Rouphael N, Edupuganti S, Weiskopf D, Tse LV, Hou YJ, Margolis D, Sette A, Collins MH, Schmitz J, Baric RS, de Silva AM. The Receptor Binding Domain of the Viral Spike Protein Is an Immunodominant and Highly Specific Target of Antibodies in Sars-Cov-2 Patients. Sci Immunol. 2020;5(48). doi: 10.1126/sciimmunol.abc8413. PubMed PMID: 32527802; PMCID: PMC7292505.
27. Food and Drug Administration. Convalescent Plasma Emergency Use Authorization Letter In: Services USDoHaH, editor. 2021.
28. Software G. Graphpad Prism. San Diego, California USA: GrapPad Software.
29. Nickel O, Rockstroh A, Borte S, Wolf J. Evaluation of Simple Lateral Flow Immunoassays for Detection of Sars-Cov-2 Neutralizing Antibodies. Vaccines (Basel). 2022;10(3). doi: 10.3390/vaccines10030347. PubMed PMID: 35334979; PMCID: PMC8949379.
30. Noval MG, Kaczmarek ME, Koide A, Rodriguez-Rodriguez BA, Louie P, Tada T, Hattori T, Panchenko T, Romero LA, Teng KW, Bazley A, de Vries M, Samanovic MI, Weiser JN, Aifantis I, Cangiarella J, Mulligan MJ, Desvignes L, Dittmann M, Landau NR, Aguero-Rosenfeld M, Koide S, Stapleford KA. Antibody Isotype Diversity against Sars-Cov-2 Is Associated with Differential Serum Neutralization Capacities. Sci Rep. 2021;11(1):5538. doi: 10.1038/s41598-021-84913-3. PubMed PMID: 33692390; PMCID: PMC7946906.
31. Biodefense G. H5n1 on the Brink: Global Virologists Urge Immediate Action to Thwart a Potential Pandemic: Global Biodefense; 2025 [08/14/2025]. Available from: https://globalbiodefense.com/2025/05/03/h5n1-on-the-brink-global-virologists-urge-immediate-action-to-thwart-a-potential-pandemic/.
32. Feng S, Phillips DJ, White T, Sayal H, Aley PK, Bibi S, Dold C, Fuskova M, Gilbert SC, Hirsch I, Humphries HE, Jepson B, Kelly EJ, Plested E, Shoemaker K, Thomas KM, Vekemans J, Villafana TL, Lambe T, Pollard AJ, Voysey M, Oxford CVTG. Correlates of Protection against Symptomatic and Asymptomatic Sars-Cov-2 Infection. Nat Med. 2021;27(11):2032-40. doi: 10.1038/s41591-021-01540-1. PubMed PMID: 34588689; PMCID: PMC8604724.
33. Fulford TS, Van H, Gherardin NA, Zheng S, Ciula M, Drummer HE, Redmond S, Tan HX, Boo I, Center RJ, Li F, Grimley SL, Wines BD, Nguyen THO, Mordant FL, Ellenberg P, Rowntree LC, Kedzierski L, Cheng AC, Doolan DL, Matthews G, Bond K, Hogarth PM, McQuilten Z, Subbarao K, Kedzierska K, Juno JA, Wheatley AK, Kent SJ, Williamson DA, Purcell DFJ, Anderson DA, Godfrey DI. A Point-of-Care Lateral Flow Assay for Neutralising Antibodies against Sars-Cov-2. EBioMedicine. 2021;74:103729. doi: 10.1016/j.ebiom.2021.103729. PubMed PMID: 34871960; PMCID: PMC8641961.
34. Lake DF, Roeder AJ, Kaleta E, Jasbi P, Pfeffer K, Koelbela C, Periasamy S, Kuzmina N, Bukreyev A, Grys TE, Wu L, Mills JR, McAulay K, Gonzalez-Moa M, Seit-Nebi A, Svarovsky S. Development of a Rapid Point-of-Care Test That Measures Neutralizing Antibodies to Sars-Cov-2. J Clin Virol. 2021;145:105024. doi: 10.1016/j.jcv.2021.105024. PubMed PMID: 34781240; PMCID: PMC8567411.
35. Suntronwong N, Assawakosri S, Kanokudom S, Yorsaeng R, Auphimai C, Thongmee T, Vichaiwattana P, Duangchinda T, Chantima W, Pakchotanon P, Chansaenroj J, Nilyanimit P, Srimuan D, Thatsanatorn T, Sudhinaraset N, Wanlapakorn N, Mongkolsapaya J, Poovorawan Y. Strong Correlations between the Binding Antibodies against Wild-Type and Neutralizing Antibodies against Omicron Ba.1 and Ba.2 Variants of Sars-Cov-2 in Individuals Following Booster (Third-Dose) Vaccination. Diagnostics (Basel). 2022;12(8). doi: 10.3390/diagnostics12081781. PubMed PMID: 35892491; PMCID: PMC9394243.
36. Zambrana JV, Mellis IA, Shotwell A, Maier HE, Saborio Y, Barillas C, Lopez R, Vasquez G, Plazaola M, Sanchez N, Ojeda S, Gilbertson I, Kuan G, Wang Q, Liu L, Balmaseda A, Ho DD, Gordon A. Variant-Specific Antibody Correlates of Protection against Sars-Cov-2 Omicron Symptomatic and Overall Infections. Nat Commun. 2025;16(1):10305. doi: 10.1038/s41467-025-65235-8. PubMed PMID: 41271716; PMCID: PMC12639126.
37. Zhong D, Xiao S, Debes AK, Egbert ER, Caturegli P, Colantuoni E, Milstone AM. Durability of Antibody Levels after Vaccination with Mrna Sars-Cov-2 Vaccine in Individuals with or without Prior Infection. JAMA. 2021;326(24):2524-6. doi: 10.1001/jama.2021.19996. PubMed PMID: 34724529; PMCID: PMC8561429.
38. Williams WW, Ingelfinger JR. Third Time’s a Charm - Covid-19 Vaccine Hope for Solid-Organ Transplant Recipients. N Engl J Med. 2021;385(13):1233-4. doi: 10.1056/NEJMe2112866. PubMed PMID: 34379913; PMCID: PMC8385552.
39. Kamar N, Abravanel F, Marion O, Couat C, Izopet J, Del Bello A. Three Doses of an Mrna Covid-19 Vaccine in Solid-Organ Transplant Recipients. N Engl J Med. 2021;385(7):661-2. doi: 10.1056/NEJMc2108861. PubMed PMID: 34161700; PMCID: PMC8262620.
40. Staying up to Date with Covid-19 Vaccines: Center for Disease Control and Prevention; 2025 [updated 6/6/2025; cited 2025 10/2/2025]. Available from: https://www.cdc.gov/covid/vaccines/stay-up-to-date.html.
Submitted August 27, 2025 | Accepted November 24, 2025 | Published February 16, 2026
Copyright © 2026 The Authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International License.